Taxonomy: Kingdom - Plantae (plants). Subkingdom - Tracheobionta (vascular plants). Superdivision - Spermatophyta (seed plants). Division - Coniferophyta (conifers). Class - Pinopsida. Order - Pinales. Family - Pinaceae (pines). Genus - Abies Mill. Species - Abies magnifica A. Murray bis
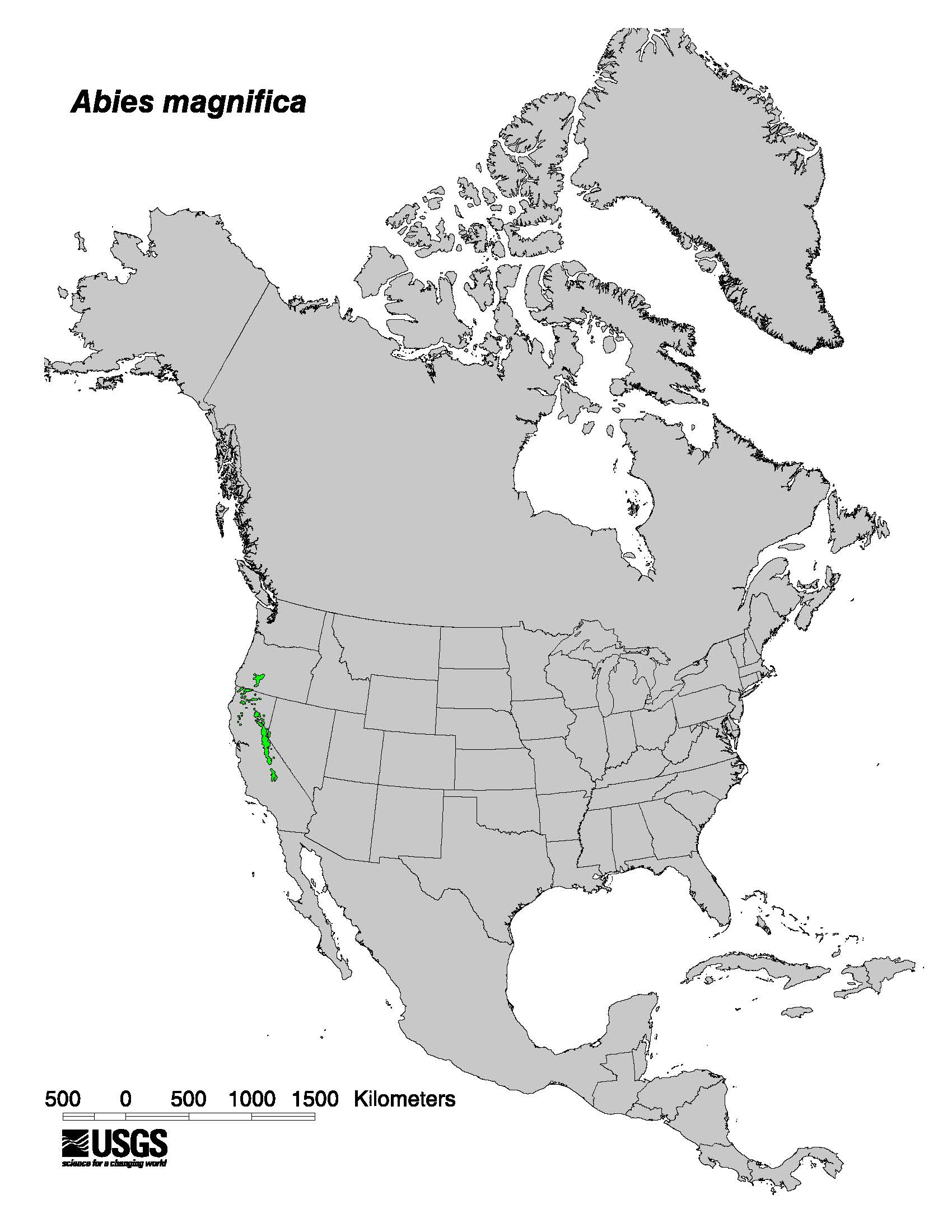
Abies magnifica often exists in extensive high elevation stands in the Sierra Nevada; its close relative A . procera occurs in small mountaintop populations relatively isolated from one another. As expected for isolated populations, A . procera produces large interpopulation variation in morphology (J.Maze and W.H. Parker 1983) and chemistry (E.Zavarin et al. 1978). Where the two species meet in southern Oregon and northern California, many populations are intermediate; these have been called A . magnifica var. shastensis Lemmon. The status of such intermediates is unsettled. They may be accepted as hybrids between A . magnifica and A . procera (Liu T. S. 1971) or, alternatively, the paleontological record suggests that the two species may have originated from the intermediates (E.Zavarin et al. 1978). Individuals from this region should be assigned to A . magnifica , A . procera , or A . magnifica × procera (E.L. Parker 1963), depending on the morphologic criteria selected to differentiate the species, though clearly these individuals are genetically quite different from those near the type localities of the two species. California red fir hybridizes with noble fir (A. procera) where they occur together. These hybrids are similar to Shasta red fir, which increases taxonomic confusion of the California red fir-noble fir complex in the Klamath region. Morphological comparisions, artifical crosses, and molecular studies indicate that Shasta red fir resulted from California red fir and noble fir introgression.Ecology: California red fir occurs in pure, dense forests between the lower montane white fir (Abies concolor) or mixed-conifer forests and the upper montane or subalpine lodgepole pine (Pinus contorta var. murrayana) and mountain hemlock (Tsuga mertensiana) forests. In the upper montane coniferous forests, California red fir is an overstory dominant on mesic sites. Canopies can be open or closed, and understory vegetation is variable but generally sparse.